

TEL:17312606166(魏經理)



美鳳力臨床前大動物實驗中心

 17312606166
17312606166
最近氣溫驟降,北方已經開始飄雪,南方也是秋雨連綿,有的小伙伴把自己包裹的嚴嚴實實,仍然難以抵抗秋季流感大軍;看著別人精神抖擻,穿個短袖到處溜達,我等凡夫俗子只能感嘆自己免疫系統的戰斗力太渣。實際上在人漫長的一生中,免疫系統都在盡職盡責的守護著大家的健康,大多數疾病都是由于免疫系統故障所引起的,如果沒有免疫系統的保護,即使是一粒塵埃都足以致命......巨噬細胞消滅病原體(引自工作細胞)
今天小編就為大家詳細介紹一下免疫系統,全文較長,大家可根據目錄瀏覽自己感興趣的部分。
01 免疫系統概述 免疫系統的分類、免疫器官的分類、免疫細胞的起源和分類、免疫活性物質
02 免疫細胞的功能 DC細胞、巨噬細胞、單核細胞、NK細胞、粒細胞、T細胞、B細胞、常用研究模型
03 免疫研究熱點及常用模型 細胞治療、免疫檢查點、常用研究模型
免疫系統由免疫器官、免疫細胞和免疫活性物質組成,具有免疫監視、防御、調控的作用,是機體執行免疫應答及免疫功能的重要系統。免疫系統具有識別和排除抗原性異物、與機體其他系統相互協調,共同維持機體內環境穩定和生理平衡的功能[1],但其功能的亢進也會對自身器官或組織產生傷害。
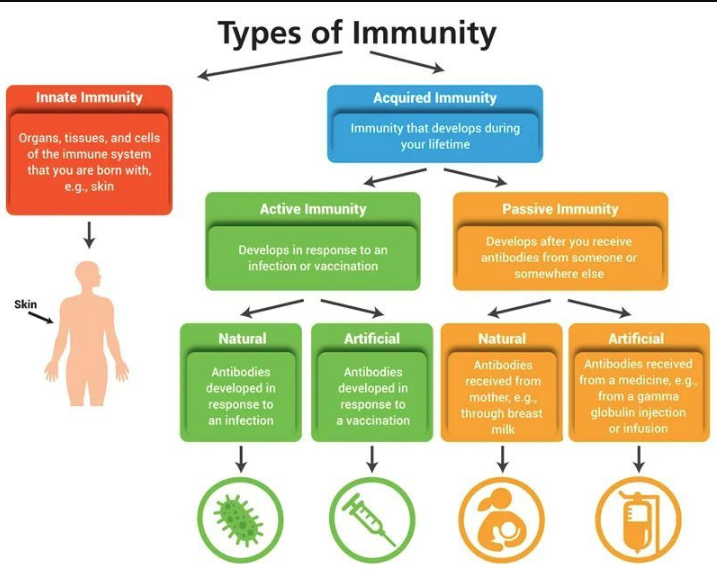
免疫系統可分為先天性免疫(又稱非特異性免疫)和適應性免疫(又稱特異性免疫),其中適應性免疫又分為體液免疫和細胞免疫,他們構筑了三道防線守護著我們的健康。第一道防線是由皮膚和黏膜構成的,它們不僅能夠阻擋病原體侵入人體,而且它們的分泌物(如乳酸、脂肪酸、胃酸和酶等)還有殺菌的作用;第二道防線是體液中的殺菌物質和吞噬細胞;第三道防線主要由免疫器官(胸腺、淋巴結和脾臟等)和免疫細胞(淋巴細胞)組成。
免疫系統分類
前兩道防線是人類在進化過程中逐漸建立起來的天然防御功能,特點是人人生來就有,不針對某一種特定的病原體,對多種病原體都有防御作用,因此屬于非特異性免疫。第三道防線是人體在出生以后逐漸建立起來的后天防御功能,只針對某一特定的病原體或異物起作用,因而屬于特異性免疫。接種疫苗、感染病原體、體外輸入抗體等方法均可讓我們獲得針對相應病原體的特異性免疫力。
免疫器官根據分化的早晚和功能不同,可分為中樞免疫器官和外周免疫器官。中樞免疫器官包括骨髓和胸腺,是免疫細胞發生、分化、成熟的場所。其中,骨髓富含多能造血干細胞(HSC),是各種血細胞的重要發源地。胸腺是T細胞分化、成熟的場所,對外周免疫器官和免疫細胞具有重要的調節作用,但成年后,胸腺會隨著年齡增長逐漸萎縮,到老年時基本被脂肪組織所取代,這也導致人的免疫力隨之下降[1]。人體免疫器官
外周免疫器官包括脾臟、淋巴結、扁桃體、小腸集合淋巴結、盲腸等,是T、B淋巴細胞定居、增殖的場所及發生免疫應答的主要部位[2]。脾臟承擔著過濾血液的職能,它可以清除死亡的血細胞,吞噬血液中的病毒和細菌;淋巴結擁有數十億個白細胞,是免疫應答的重要場所,肩負著過濾淋巴液的工作;盲腸和扁桃體內含有大量的淋巴結,割除扁桃體后人患上鏈球菌咽喉炎等疾病的幾率明顯升高......
大部分免疫細胞來源于骨髓,骨髓里的HSC可分化為不同的造血祖細胞,進而分化為形態和功能不同的髓系祖細胞(CMP)和淋巴系祖細胞(CLP)。CLP可進一步分化成T細胞、B細胞等;CMP則可以進一步分化為粒細胞、巨噬細胞、紅細胞、血小板等;CLP和CMP皆參與樹突狀細胞(DC)的形成[3]。HSC的分化路徑[8]
免疫細胞的種類繁多,大致也可分為2類。先天免疫系統的組成細胞:DC細胞、NK細胞、巨噬細胞、樹突狀細胞、肥大細胞、嗜酸性粒細胞、嗜堿性粒細胞等;適應性免疫應答細胞:T細胞、B細胞等。
免疫活性物質指由免疫細胞或其它細胞產生的發揮免疫作用的物質,免疫活性物質的種類非常多,包括抗體、溶菌酶、補體、免疫球蛋白、干擾素、白細胞介素、腫瘤壞死因子等[7]。通過免疫學檢測技術,可以對這些免疫活性物質進行定性定量分析,從而對人體自身免疫情況、營養情況、感染情況及自身免疫性疾病進行診斷。
免疫細胞是人體執行免疫功能的“一線干警”,向來是免疫研究的重點,也是我們要重點介紹的部分。
樹突狀細胞(dendritic cell, DC),存在于血液和暴露于環境的組織中,如皮膚和鼻子、肺、胃和小腸的上皮組織。它是一種專業的抗原提呈細胞,通過對T、B淋巴細胞的抗原加工和提呈,將先天免疫和適應性免疫聯系起來[11,12]。樹突狀細胞的常規標記物包括CD11c、BDCA-1/2以及CD123[13],樹突狀細胞主要分為三類:
漿細胞樣樹突狀細胞(pDC),專精于識別病毒和腫瘤細胞
I型經典樹突狀細胞(cDC1),能夠識別細胞內病原體并觸發CD8 T 細胞和Th1 CD4 T細胞應答
II型經典樹突狀細胞(cDC2),在與細胞內病原體、寄生蟲、變應原、真菌和細胞外細菌接觸時,會觸發CD4 T細胞反應
小鼠樹突細胞發育譜系圖
巨噬細胞(Macrophages)是一種分布廣泛且功能多樣的白細胞,作為“清道夫”,巨噬細胞可通過吞噬細胞殘片、細胞代謝物,消化病原體,清除有害物質;作為“哨兵”,巨噬細胞可通過細胞因子等信號彈提醒其它免疫細胞“有敵入侵,準備戰斗”;此外,巨噬細胞還可以感受微環境的變化,響應器官需求,維持代謝平衡[14]。巨噬細胞的來源[16]
巨噬細胞存在于人體的多種組織中,如骨巨噬細胞(破骨細胞)、中樞神經系統(小膠質細胞)、肝臟(庫普弗細胞),肺泡(噬塵細胞)等,不同組織的巨噬細胞具有不同的形態和功能。這些駐留在組織中的巨噬細胞可以從骨髓中的造血干細胞發育而來,它們表達F4/80,并在成熟階段發育為Ly6C+單核細胞;也可以是胚胎發育期的胎肝、卵黃囊或背主動脈產生,這些巨噬細胞在胎兒出生前就遷移到特定的組織中,并具有自我更新的能力[15],如小膠質細胞和庫普弗細胞能夠在白細胞介素34 (IL-34)存在的情況下實現自我更新;此外,髓外造血也能產生一些巨噬細胞,例如脾臟。
巨噬細胞極化[18]
根據巨噬細胞在炎癥反應中的功能和活化程度可分為兩個亞型:抗炎的M1巨噬細胞和促炎的M2巨噬細胞。M1型巨噬細胞通常由病原體、LPS、粒細胞巨噬細胞集落刺激因子(GM-CSF)、腫瘤壞死因子α(TNF-α)和1型輔助性T (Th1)細胞因子、干擾素γ(IFN-γ)等活化,它可以促進氧化狀態、產生活性氧(ROS),并分泌炎癥細胞因子和趨化因子,如IL-1、IL-6、IL-12、TNF-α、CXCL9等[17]。M2型巨噬細胞被認為具有免疫抑制作用,可由寄生蟲或真菌感染、免疫復合物、凋亡細胞、巨噬細胞集落刺激因子(M-CSF)、IL-13、TGF-b和2型輔助性T細胞因子 (Th2)IL-4、IL-33和IL-25通過Th2細胞替代活化,它可以分泌IL-10和TGF-β等抗炎因子,M2型巨噬細胞又可分為M2a、M2b、M2c及M2d四個亞型。
單核細胞(monocytes)是體積最大的白細胞,來源于骨髓中的造血干細胞,在維持機體穩態、病原體識別和清除以及炎癥中發揮關鍵作用。單核細胞在穩態下不增殖,一旦機體受到病原體攻擊,單核細胞就會迅速遷移到外周組織,分化為樹突狀細胞或巨噬細胞。單核細胞的特征是可塑性和異質性,因為它可以快速調整其功能表型以響應變化的機體環境[22]。
單核細胞的發育譜系[21]
(CDP,樹突狀細胞共同前體;CMoP,單核細胞共同祖細胞)
根據CD14、CD16 (Fcγ RIII)、CD64 (Fcγ RI)和趨化因子受體CD192和CX3CR1的表達水平可將人外周血單核細胞群定義為三個不同的亞群:經典型、中間型和非經典型;根據HLA-DR、CD195、TNFR1(CD120a)和TNFR2(CD120b)的表達水平可以對這些亞群進一步細分。TNFR1在中間型單核細胞中表達最高,而TNFR2在非經典型單核細胞中表達最高[23]。
| 亞群 | 標志物 | 功能 |
| 經典型 | CD14Hi、CD64、CD62L 、TNFR1、TNFR2Low | 占比87-88%,有吞噬活性的主要群體,較弱促炎細胞因子生成 |
| 中間型 | CD16、CD14Hi、CD64、HLA-DRHi、TNFR1Hi、TNFR2 | 占比約2-3%,促炎,生成TNF-α、IL1b和IL-6 |
| 非經典型 | CD14Low、CD16Hi、 TNFR1Low、TNFR2Hi | 占比約10%,抗炎,組成型生成IL-1RA |
(Low:低表達水平,Hi:高表達水平)
自然殺傷細胞(natural killer cell,NK)來源于骨髓CD34+共淋巴祖細胞,是先天免疫系統的一部分,對于免疫監視和隨后的宿主防御病毒感染和癌細胞至關重要。在人體中,NK細胞占循環淋巴細胞的8-20%;在實驗室的近交系小鼠中,NK細胞占脾臟和骨髓淋巴細胞的2-5%[5]。與其他淋巴細胞不同,NK細胞無需事先活化,就可以非特異性直接殺傷腫瘤細胞和病原體,這種天然殺傷活性既不需要抗原致敏,也不需要抗體參與,且無MHC限制。也因此,NK細胞不會生成免疫記憶或產生長期保護性免疫[9]。
NK細胞亞群
在人體內NK細胞主要特征為CD3-CD56+淋巴細胞群,NK細胞根據CD56的表達水平被進一步區分:dim和bright。CD56dim是一種完全成熟的NK細胞,占外周血NK細胞的90%,主要起介導細胞毒性的作用,具有更強的殺傷活性[10];CD56bright可產生大量細胞因子,主要起免疫調節作用,高表達IL-2R。除上述2種常見的NK細胞外,還有組織駐留NK細胞(trNK),它們存在于許多組織中,包括淋巴結、胸腺、肝臟、肺、子宮和小腸,我們對trNK的了解仍處于早期階段;另外一種不太常見的亞群,稱為適應性NK細胞,它們具有免疫記憶,是特化的記憶樣細胞。
粒細胞(granulocytes)是富含細胞質顆粒的細胞,一旦被活化,就會釋放免疫刺激分子來抵抗病毒和寄生蟲感染。粒細胞依靠炎癥信號將其募集到損傷、感染或過敏反應的部位,除了能對病毒和寄生蟲感染反應之外,粒細胞還與多種疾病相關,包括慢性炎癥、哮喘、過敏、免疫調節、自身免疫和癌癥等[24]。
粒細胞分類
粒細胞包括嗜中性粒細胞(neutrophils)、嗜酸性粒細胞(eosinophils)、嗜堿性粒細胞(basophils)及肥大細胞(mast cells),其標志物和功能簡介如下:
| 亞群 | 標志物 | 功能 |
| 中性粒細胞 | CD10、CD15、CD17、CD24、CD35、CD43、CD66、CD89、CD282 (TLR2)、CD284 (TLR4)、CD286 (TLR6)、鈣衛蛋白(S100A8/A9)等 | 占人體循環白細胞的60%,中性粒細胞的主要功能是吞噬病原體,消滅微生物 |
| 嗜酸性粒細 | CD9、CD15、CD24、CD35、CD125、CD170(SiglecF)、CD193 (CCR3)等 | 約占循環白細胞的1-3%,主要參與人體對寄生蟲感染和過敏性疾病的反應 |
| 嗜堿性粒細胞 | CD9、CD11a、CD13、CD16、CD25、CD88、CD154、CD123、FceR1、CD192 (CCR2)等 | 在循環白細胞中占比不到1%,是唯一含有組胺并分泌包括IL-4在內的某些細胞因子的循環白細胞,主要作用是防御寄生蟲感染 |
| 肥大細胞 | CD117 (c-kit)、FceR1、CD9、CD15、CD24、CD35、CD43、CD64等 | 廣泛分布在皮膚及內臟粘膜下的微血管周圍,分泌多種細胞因子,參與免疫調節(TB細胞,APC細胞活化 |
T細胞(T lymphocyte)起源于骨髓,然后轉移到胸腺中發育成熟,成熟的T細胞經淋巴管、外周血和組織液等在體內進行循環,T細胞的循環有利于廣泛接觸抗原物質,加強免疫應答,形成并長期保持免疫記憶。在過去的十年中,T細胞一直是研究熱點,例如研究T細胞受體的免疫檢查點抑制劑(PD-1、CTLA-4等)。目前已經有方法可以解除腫瘤微環境對T細胞增殖和激活的抑制,這一成果已經成功應用于黑色素瘤以及其他腫瘤的治療。
T細胞分化
T細胞是相當復雜的不均一體、又不斷在體內更新、在同一時間可以存在不同發育階段或功能的亞群。T細胞的特征標記物包括CD3和T細胞受體(TCR)。根據表達類型,它們可以分為CD4+ T細胞和CD8+ T細胞,其中CD4+ T細胞主要為輔助T細胞(helper T cell),它可以增生擴散來激活其它直接參與免疫反應的免疫細胞;CD8+ T細胞主要為細胞毒T細胞(cytotoxic T cell),它就像一個“殺手”,可以對產生特殊抗原反應的目標細胞進行直接殺滅;還有一種調節性T細胞(T-reg),它能有效抑制免疫應答,通常起著維持自身耐受和避免免疫反應過度損傷機體的重要作用,目前研究最多的是CD4+CD25+ Treg細胞(nTreg),此外還有CD+8 Treg、aTreg、iTreg等;此外還有記憶T細胞、NKT細胞等。CD4+ T細胞和CD8+ T細胞根據表面標志物又可以進一步分為更多亞群:
CD4+和CD8+T細胞亞群
| 亞群 | 標志物 | 功能 |
| 輔助性T細胞1(Th1) | CD3、CD4、IFN-g、CCR5、CXCR3等 | Th1細胞在識別和清除細胞內病原體如病毒和細菌,包括結核分枝桿菌、麻風分枝桿菌和利什曼原蟲方面起著重要作用。 |
輔助性T細胞2(Th2) | CD3、CD4、CCR3、CCR4、CXCR4、IL-4、IL-10等 | Th2細胞介導針對細胞外寄生蟲、細菌、過敏原和毒素的體液或抗體介導的免疫反應的活化和維持。Th2細胞通過生成各種細胞因子如IL-4、IL-5、IL-6、IL-9、IL-13和IL-17E (IL-25)來介導這些功能。 |
| 輔助性T 胞17(Th17) | CD3、CD4、CCR4、CCR6、TGFbRII、IL-17、CCL20等 | 一種新發現的能夠分泌白介素17(IL-17)的T細胞亞群,在自身免疫性疾病和機體防御反應中具有重要的意義。 |
| 調節性T細胞(T-reg) | CD3、CD4、FoxP3、CD25、CTLA4、OX40/CD134、TGFb、IL-10、IL-35等 | Treg對維持自身耐受性和免疫細胞穩態至關重要,Treg在調節自身免疫性疾病的免疫反應中發揮了重要作用,包括1型糖尿病、類風濕性關節炎、多發性硬化等。 |
濾泡輔助T細胞(Tfh) | CD3、CD4、CD8、CD185 (CXCR5)、CD183 (CXCR3)、CD278 (ICOS)、CD279 (PD1)等 | Tfh細胞是增強免疫應答的關鍵,作用是觸發B細胞分化成分泌抗體的漿細胞和記憶性B細胞,了解它們的功能有助于對疫苗的開發。 |
| NaiveT細胞 | CD3、CD8、CD27、CD45RA、CD62L、CD127、CCR7等 | 表現靜息表型,通過TCR-自身肽、MHC配體和IL-7信號傳導維持 |
| 效應T細胞 | CD3、CD8、CD25、CD69、KLRG1、CD30、OX40、ICOS、TIM3等 | 對轉化細胞和病毒感染細胞具有細胞毒性,通過Fas/FasL和分泌IFNγ、顆粒酶A和穿孔素介導細胞死亡 |
| 效應記憶性T細胞(Tem) | CD3、CD8、CD44、CD45RO、CD127等 | 存在于淋巴組織和外周組織中;細胞毒性強和儲備好的效應分子,在遇到抗原時迅速分化為Teff |
| 中樞記憶性T細胞(Tcm) | CD3、CD8、CD45RO、CD127、CD27、CD28等 | 存在于淋巴結、脾臟、骨髓和血液;相比于naive CD8+T細胞,對抗原刺激更為敏感,但無即時效應反應 |
B細胞(B lymphocyte)在骨髓中開始發育成熟,是體內產生抗體(免疫球蛋白Ig)的細胞,主要執行體液免疫,也具有抗原提呈功能。與其他淋巴細胞一樣,B細胞的標記物也在成熟和分化過程中發生進化,包括CD19、CD27、BCM和CXCR4等。B細胞的典型特征在于B細胞受體(BCR),BCR是一種錨定在B細胞膜上的抗體,可通過細胞內信號傳導刺激B細胞的活化。每個成熟B細胞只生成一種具有單一特異性的BCR,因此只生成具有單一特異性的抗體。
B細胞分化
從骨髓來的干細胞或前B細胞,在遷入法氏囊或類囊器官后,逐步分化為有免疫潛能的B細胞。成熟的B細胞經外周血遷出,進入脾臟、淋巴結,主要分布于脾小結、脾索及淋巴小結、淋巴索及消化道粘膜下的淋巴小結中,受抗原刺激后,一部分分化增殖為漿細胞,合成抗體,發揮體液免疫的功能;另一部分B細胞經過抗原激活后并不成為漿細胞,而是成為記憶B細胞,當再次遇到相同抗原時,記憶B細胞能迅速作出反應,大量分化增殖。
以上只是簡單介紹了免疫細胞的分類,隨著研究的深入,科學家們發現了更多的Marker,分出了更多的亞群。目前免疫學的研究炙手可熱,各種顛覆性的成果不斷產生,工欲善其事必先利其器,想要在免疫學研究中搶占先機,合適的動物模型是必不可少的。
(圖片來自網絡)
南模生物根據免疫細胞發育分化的標志性marker,自主構建了多種Cre工具鼠、搭配特定基因條件性敲除小鼠、熒光工具鼠等使用,可用于研究該基因在免疫細胞亞群中的功能、或研究免疫細胞的發育分譜系;此外還有多種DTR工具鼠,可以在小鼠體內剔除特定類型的免疫細胞,如Lyz2-DTR,可以剔除小鼠體內的巨噬細胞,Tmem119-DTR,可以剔除小鼠體內的小膠質細胞等。詳細模型信息見下表:
T、B、NK細胞Cre工具鼠
造血干細胞、巨噬細胞、單核細胞等Cre工具鼠
粒細胞、DC細胞等Cre工具鼠
各類免疫細胞剔除(DTR)工具鼠
免疫系統是人類健康忠誠的衛士,90%以上的疾病都和免疫系統失調有關,如各類腫瘤、病毒感染(流感、HIV、登革熱等)、細菌感染(結核分枝桿菌、金黃色葡萄球菌等)、自身免疫性疾病(類風濕關節炎、系統性紅斑狼瘡等)等。這其中最火熱的無疑是免疫在腫瘤治療領域的研究了,我們就以腫瘤治療為例,介紹下當下的免疫研究熱點和常用動物模型。
細胞治療是指將同種或異種活細胞在體外經過加工或處理后,再輸注回人體,以達到修復病變細胞或重建正常細胞和組織的治療方法。細胞治療在心腦血管疾病、惡性腫瘤、遺傳病、自身免疫性疾病、損傷再生醫學等領域,都有非常廣闊的前景。目前,細胞治療主要還處于基礎研究階段,只有少數產品已經走上了臨床,如CAR-T等。
CAR-T療法原理圖
CAR-T即嵌合抗原受體T細胞免疫療法,原理包含四部分:
從患者身上分離T細胞
通過基因工程技術改造T細胞,使T細胞表達能識別腫瘤細胞的人工受體(CAR),改造后的T細胞帶有CAR受體(即CAR-T細胞),可以識別并攻擊癌細胞
體外培養擴增CAR-T細胞,一般需要幾十億-幾百億有效的CAR-T細胞
將擴增好的CAR-T細胞回輸到病人體內,高效滅殺癌細胞
細胞治療是一個非常前沿的領域,2017年8月,首個CAR-T免疫療法在美國上市,今年6月,復星凱特CAR-T細胞治療產品正式獲批,我國才有了自己的CAR-T產品。截至目前,全球也已有7款CAR-T產品上市,CAR-NK等則還完全停留在研究階段。
已上市CAR-T產品(至2021.9月)
當然,細胞治療自身也有著明顯的缺點:
價格太貴是細胞療法推廣的最大障礙,細胞治療需要取材于患者自身,無法像傳統藥物一樣大規模商業化生產。作為擁有首個獲批CAR-T療法的企業,諾華的CAR-T細胞療法每療程定價47.5萬美元,令人望而卻步。
細胞一般來自于腫瘤患者自身,只能個性化制備,這就為細胞的質控帶來了極大困難,有的患者因為病情原因,身上可取的T細胞并不足量或者質量較差。
腫瘤細胞一般和正常細胞表達相同的抗原,唯一區別在于某些抗原在腫瘤細胞表面高表達而在正常細胞表面低表達。根據競爭原則,CAR-T細胞會優先殺傷腫瘤細胞,但也會誤傷正常細胞。因此,發掘腫瘤細胞專屬抗原是免疫細胞治療中的一項重要工作。
每一項技術的興起和完善都是一個漫長的過程,目前細胞治療不僅本身有諸多缺陷,還面臨著來自傳統藥物、新興的免疫檢查點抑制劑等的跨界競爭,但不可否認的是,細胞治療的前景是光明的,這種讓人耳目一新的治療方法必將在未來的醫藥格局中占有重要地位。
PD-1/PD-L1的巨大成功,讓全球進入免疫檢查點抑制劑開發的大時代,每個人都不愿意被時代拋下。免疫檢查點(immune checkpoint)指的是在免疫細胞上表達,能調節免疫激活程度的一系列分子。人的免疫系統在受到激發時會活化,消滅病原體,但不會攻擊自身正常細胞,這就是因為免疫檢查點起到了剎車作用,它可以將免疫系統的活化程度控制在正常范圍內。但是免疫檢查點也會被一些“不法分子”利用,例如腫瘤細胞,它們可以釋放一些物質來激活免疫檢查點,抑制免疫系統活化,使自己逃脫免疫監視,逍遙法外。
部分常見免疫檢查點[25]
因此開發免疫檢查點抑制劑,激活免疫細胞重新工作從而消滅腫瘤細胞等“不法分子”,成為了當下炙手可熱的研究方向。免疫檢查點分子據功能區同源性可以分為幾大家族:B7-CD28超家族,免疫球蛋白(Ig)超家族和腫瘤壞死因子(TNF)超家族等,跟據其對細胞的調節作用又可分為共刺激分子與共抑制分子兩大類。部分常見免疫檢查點簡介見下表:
| 免疫檢查點 | 簡介 |
| PD-1(CD279) | PD-1在CD4和CD8胸腺細胞以及活化的T和B淋巴細胞和骨髓細胞上短暫表達。PD-1在結合B7家族的兩個配體PD-L1和PD-L2之一后發出信號。在配體結合后,PD-1信號傳導抑制T細胞活化,導致增殖減少,細胞因子產生和T細胞死亡。 |
PD-L1(B7-H1) | PD-L1在T淋巴細胞,B淋巴細胞,NK細胞,樹突細胞以及IFNγ刺激的單核細胞,上皮細胞和內皮細胞上表達。PD-L1與其受體PD-1結合,在CD4和CD8胸腺細胞以及活化的T和B淋巴細胞和骨髓細胞上發現。PD-L1與PD-1的結合導致TCR介導的T細胞增殖和細胞因子產生的抑制 |
LAG-3 | 選擇性表達在活化的T淋巴細胞、NK細胞和樹突狀細胞上。LAG-3主要的配體是MHC II類分子,對T細胞具有負調節功能 |
TIM-3 | 表達于Th1、Th17、CD8+T細胞上,是T細胞抑制受體。Gal-9可以與TIM-3結合并引起鈣流入Th1細胞的細胞內區域,誘導T細胞凋亡 |
4-1BB | 主要表達于活化的T細胞。與4-1BBL結合時,可刺激T 細胞( 和B 細胞) 活化和增殖。其介導的信號傳導可保護T細胞,特別是CD8+ T細胞免于激活誘導的細胞死亡。 |
OX-40 | OX40是活化的細胞毒性T細胞和調節性T細胞(Treg細胞)表面表達的共刺激分子。OX40與配體OX40L(CD252, TNFSF4)結合傳遞共刺激信號。在腫瘤微環境中,免疫激活可導致OX40表達。可增強效應T細胞的活化和增殖,并抑制Tregs,從而導致復雜的抗腫瘤免疫反應。 |
| TIGHT | TIGIT 在活化和記憶性 T 細胞、調節性 T 細胞、濾泡輔助性 T(Tfh)細胞和自然殺傷性(NK)細胞上表達。與 PD-1 相似,在慢性感染和腫瘤中,高表達于耗竭的 T 細胞和自然殺傷(NK)細胞。 |
CTLA-4(CD152) | CTLA-4在活化的T和B淋巴細胞上表達。CTLA-4在結構上類似于CD28,并且兩種分子都與B7家族成員B7-1(CD80)和B7-2(CD86)結合。在配體結合后,CTLA-4抑制細胞介導的免疫應答。CTLA-4在誘導和/或維持免疫耐受、胸腺細胞發育和保護性免疫調節中起作用 |
CD28 | 在幾乎所有人CD4+ T細胞和約一半的所有CD8 T細胞上組成型表達。CD28與兩種配體CD80和CD86結合,在樹突細胞上表達,結合導致T細胞擴增 |
| TNFR2(CD120b) | TNFR2,是腫瘤壞死因子(TNF)受體超家族成員,TNFR2表達于抑制調節性T細胞,髓系抑制性細胞(MDSCs),內皮細胞和生長過程中的選擇神經元。TNFR2是TNF的受體之一,僅可被mTNF完全激活并觸發有效的信號轉導,行使細胞存活和免疫抑制相關功能。 |
| NKG2A | NKG2A是NKG2家族中的抑制性成員,主要表達于NK細胞和NKT細胞。HLA-E是其天然配體。NKG2A與CD94結合形成異源二聚體,與其配體結合后,激活細胞內的抑制性信號通路,抑制NK細胞活性,從而使腫瘤避免被NK細胞殺傷。 |
CD86(B7-2) | CD86由活化的T細胞和B細胞,巨噬細胞和樹突細胞表達。該配體與CD28結合,提供T細胞活化和存活以及細胞因子產生所必需的共刺激信號。另外,CD86與抑制T細胞的CTLA-4結合。 |
CD80(B7-1) | CD80由活化的B細胞和單核細胞和樹突細胞表達。該配體與CD28結合,提供T細胞活化和存活以及細胞因子產生所必需的共刺激信號。另外,CD80與抑制T細胞的CTLA-4結合。 |
ICOS(CD278) | 誘導型T細胞共刺激分子,也稱為CD278,在活化的T細胞上表達。其配體為ICOSL,主要在B細胞和樹突狀細胞上表達。 |
CD40 | 在抗原呈遞細胞(APC),如B細胞,樹突狀細胞(DC),和單核細胞以及許多非免疫細胞和各種類型的癌細胞中表達。其配體CD40L 在CD4+ T細胞表面表達。CD40-CD40L在免疫反應中對CD8+細胞毒性T淋巴細胞(CTL)功能起著至關重要的作用,是適應性免疫應答所必需的。 |
GITR | 在靜止的CD4+和CD8+ T細胞上,GITR的表達水平是非常低的,一旦接受了刺激,GITR的表達則會被提升。GITR的配體GITRL在活化抗原提呈細胞(APC)、內皮細胞和活化T細胞中均有較高水平的表達。T細胞上GITR與APC上GITRL的相互作用是雙向的: 兩種細胞都受到相互作用的影響,最終導致T細胞增殖。 |
免疫研究用小鼠分類
免疫治療靶點人源化小鼠模型,指通過基因修飾的手段將免疫治療靶點替換為相應的人源基因,使小鼠表達人源的藥物靶點(包括免疫檢查點在內),使用合適的該類人源化小鼠模型可以大幅降低研究成本,節省研究時間,同時高效地獲得可靠的實驗數據。南模生物目前擁有300多種人源化小鼠品系(包括雙靶標、三靶標、四靶標人源化小鼠),模型資源庫涵蓋目前最新最熱的免疫治療靶點,成品人源化小鼠目錄如下:
免疫缺陷模型是指一種或多種免疫系統組成成分缺陷的動物模型,隨著基因編輯技術的不斷發展,越來越多的免疫缺陷小鼠品系被研發出來,在免疫學,傳染病學,腫瘤學,干細胞生物學等研究領域中發揮著越來越重要的作用。
主要免疫缺陷小鼠的發展歷程
免疫缺陷模型的詳細介紹可以瀏覽我們的往期推文:免疫缺陷小鼠之前世今生
具體選擇哪種免疫缺陷模型,取決于自己的研究內容,是同種異體移植還是異種移植?接種的腫瘤細胞的生長特性如何?是免疫學研究還是腫瘤學研究?下面的表格也許可以給你一些參考:
| 品系 | 簡介 | 特點 |
| Nude (nu) | Foxn1基因純合突變,無毛、無胸腺,T細胞缺失;B細胞正常、NK細胞應答代償性的更功能強 | 裸鼠是快速生長的腫瘤細胞系是理想的宿主,不適合作為血源性癌癥如白血病或淋巴瘤的宿主。 |
| Scid (scid) | Prkdcscid純合突變,T、B細胞功能缺失 | 適合同種和異種腫瘤移植,特別是移植生長緩慢、原代細胞、血源性癌癥細胞。Scid小鼠隨著年齡增長,可能會產生低水平的血清免疫球蛋白Ig;scid突變對輻射敏感,不太合適需要輻照后再進行移植的實驗;人PBMC和胎兒造血細胞(HSC)的移植效率較低 |
| NOD-scid | NOD背景下Prkdcscid純合突變,T、B細胞功能缺失,NK細胞活性低,沒有溶血補體活性,骨髓發育缺陷 | 血清免疫球蛋白Ig泄漏率非常低,非常適合同種和異種腫瘤移植。自發胸腺淋巴瘤,壽命短,約為8-9個月,不適合長期的移植實驗 |
| M-NSG | NOD背景下攜帶Prkdcscid和IL2受體γ鏈(IL2rgnull)雙基因純合突變,T、B、NK細胞功能均缺失 | 可高效地植入人CD34+造血干細胞(HSC),外周血單核細胞(PBMC),病人來源異種移植物(PDX)或成體干細胞和組織,基本上可以植入人體免疫系統,是研究人體免疫功能,傳染病,糖尿病,腫瘤學和干細胞生物學的重要免疫缺陷小鼠,是目前國際公認的免疫缺陷程度最高、最適合人源細胞或組織移植的工具小鼠。 |
| CBA/N | Btk基因突變,B細胞功能缺陷,T細胞功能正常 | 可以進行骨髓移植修復,用于研究B淋巴細胞的產生、功能和異質性,較少用于腫瘤學研究 |
| Beige(bg) | Beige突變,嚴重缺乏NK細胞,欠缺巨噬細胞的抗腫瘤活性、殺傷細胞活性等 | 應用于免疫學領域。對各種致病因子較敏感,表型與人類罕見病Chediak-Higashi綜合征非常相似。 |
南模生物自主研發了M-NSG小鼠,該品系是通過基因編輯技術獲得的NOD背景下攜帶Prkdc和IL2受體γ鏈雙基因純合突變的小鼠,與NSG或NOG小鼠一樣,缺乏成熟的T、B細胞和NK細胞,是目前免疫缺陷程度最高的小鼠之一。M-NSG小鼠對人源細胞和組織免疫排斥反應低;腫瘤細胞成瘤性好,少量細胞即可成瘤;人體細胞和組織移植存活率顯著提高;適用于人類造血干細胞移植;適合用做異源細胞和組織移植的載體小鼠。點擊查看M-NSG驗證數據
免疫系統人源化小鼠則是選擇合適的免疫缺陷小鼠(如M-NSG),把人的造血細胞、淋巴細胞、某些組織等植入免疫缺陷小鼠體內,在小鼠身上模擬出人的免疫系統。免疫系統人源化模型主要分為Hu-PBMC、Hu-HSC、Hu-BLT等類型,其詳細介紹、在人類疾病中的應用、如何選擇合適的人源化模型,點擊下文即可查看。
南模生物深耕基因編輯領域,提供全方位模式生物服務,包括基因修飾成品模型供應、個性化模型定制、飼養繁育、表型分析、藥效評價等,滿足不同實驗室需求。
Reference:
1、陳淑增,楊翀,邱丹纓主編;謝德秋,田新利,萬巧鳳副主編.病原生物學與免疫學第2版
2、夏和先,齊永長主編;彭成,蔣斌,張文霞副主編;齊永長,張文霞,尚智等編.病原生物學與免疫學基礎 第3版
3、How the Immune System Works 5th - 2016
4、https://www.bdbiosciences.com/zh-cn/learn/research/immunology
5、https://www.thermofisher.cn/cn/zh/home/life-science/cell-analysis/cell-analysis-learning-center/immunology-at-work.html
6、 Pelanda, Roberta; Torres, Raul M. Central B-Cell Tolerance: Where Selection Begins.
7、Lubbers R, van Essen MF, van Kooten C, Trouw LA. Production of complement components by cells of the immune system. Clin Exp Immunol. 2017;188(2):183-194. doi:10.1111/cei.12952
8、Cold Spring Harb Perspect Med . 2013 Apr 1;3(4):a011601. doi: 10.1101/cshperspect.a011601.
9、Held W, Jeevan-Raj B, Charmoy M (2018) Transcriptional regulation of murine natural killer cell development, differentiation and maturation.Cell Mol Life Sci 75:3371–3379。
10、Kumar S (2018) Natural killer cell cytotoxicity and its regulation by inhibitory receptors.Immunology 154:383-393。
11、Collin M, Bigley V. Human dendritic cell subsets: an update. Immunology. 2018;154(1):3-20. doi:10.1111/imm.12888
12、Takenaka MC, Quintana FJ. Tolerogenic dendritic cells. Semin Immunopathol. 2017;39(2):113-120. doi:10.1007/s00281-016-0587-8
13、Steinman, Ralph M . Decisions About Dendritic Cells: Past, Present, and Future[J]. Annual Review of Immunology, 2012, 30(1):1-22.
14、Chen Y, Zhang X (2017) Pivotal regulators of tissue homeostasis and cancer: macrophages.Exp Hematol Oncol 6:23.PMID 28804688
15、Wynn TA, Chawla A, Pollard JW (2013) Macrophage biology in development, homeostasis and disease.Nature 496:445-455。PMID 23619691
16、Nature. 2013 April 25; 496(7446): 445–455. doi:10.1038/nature12034.
17、T'Jonck W, Guilliams M, Bonnardel J (2018) Niche signals and transcription factors involved in tissue-resident macrophage development.Cell Immunol 330:43–53.PMID 29463401
18、Int Immunopharmacol . 2019 May;70:459-466. doi: 10.1016/j.intimp.2019.02.050. Epub 2019 Mar 9.
19、F1000Res . 2018 Apr 6;7:429. doi: 10.12688/f1000research.13567.1. eCollection 2018.
20、https://www.thermofisher.cn/cn/zh/home/life-science/cell-analysis/cell-analysis-learning-center/immunology-at-work/b-cell-overview.html
21、Microbiol Spectr . 2016 Oct;4(5). doi: 10.1128/microbiolspec.MCHD-0015-2015.
22、Terry RL, Miller SD (2014) Molecular control of monocyte development.Cell Immunol 291:16–21.
23、Collin M, Bigley V (2016) Monocyte, macrophage, and dendritic cell development: the human perspective.Microbiol Spectr 4(5).
24、de Ruiter K, van Staveren S, Hilvering B et al.(2018) A field-applicable method for flow cytometric analysis of granulocyte activation: Cryopreservation of fixed granulocytes.Cytometry A 93:540–547.
25、Front Immunol . 2021 Jul 1;12:694055. doi: 10.3389/fimmu.2021.694055. eCollection 2021.
| 上一篇:小鼠兩性那些事兒 | 下一篇:中國實驗動物學會黨支部組織學習習近平新時代中國特色社會主義思想主題教育活動 ——參觀首鋼園 |
|
|
|
|
