

TEL:17312606166(魏經(jīng)理)



美鳳力臨床前大動物實(shí)驗(yàn)中心

 17312606166
17312606166
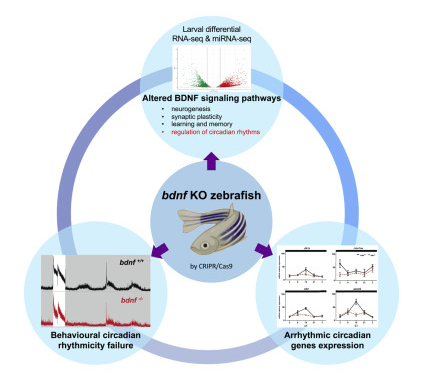
摘要:腦源性神經(jīng)營養(yǎng)因子 (BDNF) 在神經(jīng)元生長和分化、神經(jīng)元可塑性、學(xué)習(xí)和記憶中起關(guān)鍵作用。 我們使用 CRISPR/Cas9 技術(shù)構(gòu)建了一個(gè)重要的 斑馬魚Bdnf 基因敲除突變系,并對其進(jìn)行了分子和行為表征。盡管在形態(tài)學(xué)檢查中沒有發(fā)現(xiàn)明顯的缺陷,但與野生型同系胚胎相比,66% 的編碼基因和 37% 的 microRNA 在 bdnf-/- 中存在差異表達(dá)。我們深入研究了生物鐘通路,并證實(shí)了時(shí)鐘(arntl1a、clock1a 和 clock2)和時(shí)鐘控制(anaat2)基因的節(jié)律性表達(dá)的變化。通過行為測試驗(yàn)證了 Bdnf 對斑馬魚生物鐘的調(diào)節(jié)作用,突出了 bdnf-/- 仔魚缺乏晝夜活動節(jié)律。通過藥物治療可挽救部分晝夜節(jié)律行為。 這里介紹的 bdnf-/- 斑馬魚系是第一個(gè)有價(jià)值且穩(wěn)定的脊椎動物模型,用于研究 BDNF 相關(guān)的神經(jīng)發(fā)育疾病。
關(guān)鍵詞:魚仔魚:bdnf基因敲除導(dǎo)致晝夜節(jié)律失調(diào)神經(jīng)行為 分子神經(jīng)科學(xué) 發(fā)育神經(jīng)科學(xué) 細(xì)胞神經(jīng)科學(xué)
簡介: 腦源性神經(jīng)營養(yǎng)因子(BDNF)是一種小的分泌性蛋白質(zhì),屬于神經(jīng)營養(yǎng)因子家族。自發(fā)現(xiàn)以來,BDNF一直被認(rèn)為在中樞神經(jīng)系統(tǒng)發(fā)育過程中對調(diào)節(jié)離散神經(jīng)元群的分化和存活至關(guān)重要。通過結(jié)合兩類跨膜受體(泛神經(jīng)營養(yǎng)因子 p75 受體和原肌球蛋白相關(guān)激酶 (Trk) 家族受體)發(fā)揮作用,對 TrkB 亞型具有較高的親和力。與這些分子的相互作用導(dǎo)致主要信號通路的激活,這些通路調(diào)節(jié)下游基因的表達(dá),這些基因參與調(diào)節(jié)特定神經(jīng)元靶細(xì)胞的生長、維持和存活。BDNF除了在神經(jīng)系統(tǒng)發(fā)育中起到公認(rèn)的作用外,還具有在學(xué)習(xí)和記憶過程中增強(qiáng)突觸可塑性的功能。大量證據(jù)表明,BDNF 在長期增強(qiáng)中發(fā)揮關(guān)鍵作用,突觸可塑性的一種形式被廣泛認(rèn)為是研究長期記憶形成的細(xì)胞模型。BDNF 還參與調(diào)節(jié)晝夜節(jié)律。 它在嚙齒動物下丘腦的視交叉上核 (SCN) 有節(jié)奏地表達(dá),BDNF 還參與調(diào)節(jié)晝夜節(jié)律。 它在嚙齒動物下丘腦的視交叉上核 (SCN) 有節(jié)奏地表達(dá),并在光夾帶中起重要作用。迄今為止,關(guān)于BDNF作用的大部分可用信息都來自于在小鼠進(jìn)行的研究,但整個(gè)情況尚未完全闡明。主要障礙是bdnf純合子缺失的早期產(chǎn)后致死性。因此,所有結(jié)果僅限于雜合子動物,這些動物沒有表現(xiàn)出明顯的表型受損。
BDNF在脊椎動物中非常保守,哺乳動物和魚類之間的序列同源性較高(>90%)。雖然Bdnf和其他神經(jīng)營養(yǎng)因子已經(jīng)在斑馬魚中進(jìn)行了研究,但它們在魚類品系的機(jī)制層面上的研究卻很少。最近,這種神經(jīng)營養(yǎng)素在斑馬魚的大腦再生、食欲和新陳代謝中發(fā)揮了作用。與哺乳動物相比,在斑馬魚中有可能產(chǎn)生可行的敲除突變體。在這項(xiàng)研究中,我們使用 CRISPR/Cas9 技術(shù)產(chǎn)生了一個(gè)具有穩(wěn)定且可遺傳的純合 40 堿基對 (bp) 敲除的 bdnf-/- 斑馬魚系,并分析了突變體與對照的轉(zhuǎn)錄和行為表型。我們觀察到Bdnf突變體在游泳活動中表現(xiàn)出明顯的缺陷,這與Bdnf在運(yùn)動和學(xué)習(xí)等復(fù)雜行為中的既定作用一致。差異表達(dá)分析(RNA-seq 和 qPCR)揭示了與晝夜節(jié)律系統(tǒng)有關(guān)的關(guān)鍵基因表達(dá)的改變。 此外,在本研究中,我們旨在研究 bdnf 突變仔魚體內(nèi)的日常和晝夜節(jié)律活動。雖然突變體仔魚的光夾帶沒有受損,但在持續(xù)的黑暗中,節(jié)律性強(qiáng)烈減弱,表明Bdnf在晝夜節(jié)律活動中的作用。有趣的是,我們還發(fā)現(xiàn)一些參與調(diào)節(jié)時(shí)鐘基因表達(dá)的 miRNA 在突變體中表達(dá)不同。根據(jù)我們的研究結(jié)果,我們建議將 bdnf-/- 斑馬魚作為一種理想的脊椎動物模型,用于闡明 Bdnf 在復(fù)雜行為(如運(yùn)動、進(jìn)食和學(xué)習(xí))以及抑郁和焦慮相關(guān)疾病中的作用。
斑馬魚bdnf-CRISPR/Cas9基因敲除:為了研究Bdnf在斑馬魚神經(jīng)系統(tǒng)發(fā)育過程中的作用,我們構(gòu)建了一個(gè)CRISPR/Cas9介導(dǎo)的敲除系,位于影響所有五種ORF基因亞型的起始密碼子下游Bdnf第2外顯子缺失40 bp。使用T7內(nèi)切酶I(T7EI)檢測從微量注射胚胎中提取的基因組DNA,評估針對所選區(qū)域的gRNA誘變的效率。F0突變魚與野生型(wt)魚雜交。 雜合種群 (bdnf+/-) 培育一代 (F2) 以減少嵌合體和意外的脫靶突變。使用T7EI分析和Sanger測序,我們驗(yàn)證了nova2基因中沒有突變,這是CRISPR設(shè)計(jì)工具預(yù)測的唯一脫靶基因。F3 代反映了 25% bdnf+/+、50% bdnf++/- 和 25% bdnf-/- 魚的孟德爾比率。對每種情況下的胚胎池總蛋白裂解物進(jìn)行的Western印跡分析證實(shí),Bdnf+/-魚中的Bdnf蛋白水平約為Bdnf+/+對照組的一半,純合Bdnf?/?中的Bdnf蛋白水平幾乎消失。雖然我們沒有預(yù)料到完整突變體的蛋白質(zhì)裂解物中會有任何陽性信號,但我們在蛋白質(zhì)印跡中觀察到一條微弱的條帶,這可能是由于母體 Bdnf 或異源抗 BDNF 抗體與其他神經(jīng)營養(yǎng)因子的交叉反應(yīng)。

圖 1. bdnf-/- 斑馬魚系的生成
因此,我們得出結(jié)論是我們成功地建立了穩(wěn)定的、可遺傳的bdnf-/- 斑馬魚系。重要的是,在突變胚胎中未觀察到發(fā)育過程中的形態(tài)異常和致死性。雄性和雌性突變魚都能進(jìn)入成年期,并表現(xiàn)出正常的繁殖成功率。
bdnf 敲除仔魚的行為:除了已知在神經(jīng)元、存活和分化中的作用外,BDNF還在突觸可塑性的調(diào)節(jié)中發(fā)揮作用。因此,我們推測它的缺失可能會影響調(diào)節(jié) bdnf 突變體運(yùn)動活動的神經(jīng)回路。 為了驗(yàn)證這一假設(shè),我們測量了受精后 4 天 (dpf) 仔魚對光刺激的反應(yīng)。與bdnf+/+(n=32)對照仔魚相比,突變仔魚表現(xiàn)出運(yùn)動缺陷,雜合子bdnf+/-(n=32)仔魚的活性顯著降低,純合子bdnf?/? n=32)仔魚的強(qiáng)烈運(yùn)動抑制。通過三個(gè)參數(shù)來測量運(yùn)動缺陷:光刺激后10分鐘內(nèi)的總行程、速度和旋轉(zhuǎn)頻率。與 bdnf+/+ 仔魚所有三個(gè)測量參數(shù)相比,bdnf+/- 仔魚的減少并不顯著,而 bdnf-/- 仔魚影響嚴(yán)重。此外,在測試期間斑馬魚在箱內(nèi)路徑的軌跡進(jìn)行的分析表明bdnf?/? 仔魚表現(xiàn)出明顯的趨向性,,即動物傾向于避開箱的中心并趨向其邊緣或外圍移動 ,而 bdnf+/+ 和 bdnf+/- 仔魚無明顯邊緣趨向。由于趨向性是動物中一個(gè)經(jīng)過充分驗(yàn)證的焦慮指數(shù),該結(jié)果也證實(shí)了Bdnf在斑馬魚應(yīng)激相關(guān)行為中的作用。為了證明觀察到的表型與 Bdnf 的缺乏有關(guān),我們通過用 10 μM 的 7,8-二羥基黃酮水合物 (7,8-DHF)(一種合成分子)處理 22 hpf bdnf-/- 胚胎進(jìn)行了藥理學(xué)實(shí)驗(yàn) 通過激活其 TrkB 受體來模擬 Bdnf 的作用。對 7,8-DHF 處理的 bdnf-/- 突變仔魚 (n = 32) 的行為測試表明,與 bdnf-/- 仔魚 (n = 89) 相比,運(yùn)動活動的恢復(fù)具有統(tǒng)計(jì)學(xué)意義。這些結(jié)果證實(shí),Bdnf缺乏會影響斑馬魚仔魚的運(yùn)動活動和趨向性。

圖 2. bdnf+/- 和 bdnf-/- 斑馬魚仔魚的行為

圖 3. bdnf-/- 斑馬魚仔魚的藥理試驗(yàn)測試
使用 RNA-seq 對 bdnf-/- 系進(jìn)行差異基因表達(dá)分析:我們對 48 hpf bdnf-/- 突變體和 bdnf+/+ 胚胎進(jìn)行了深度 RNA 測序 (RNA-seq)。每個(gè)樣本平均產(chǎn)生 2800 萬個(gè)短配對末端片段,其中大約 89% 被明確定位到斑馬魚基因組。我們使用 DESeq2 方法在 48 hpf bdnf-/- 突變體和 bdnf+/+ 胚胎之間鑒定出 2614 個(gè)上調(diào)基因和 1653 個(gè)下調(diào)基因,表明與 bdnf突變相關(guān)的顯著轉(zhuǎn)錄差異。值得注意的是,參與神經(jīng)營養(yǎng)途徑的關(guān)鍵基因的不同表達(dá)水平的上調(diào),例如Trk2a(主要Bdnf和Nt4/5受體)、Trk3a(Nt3受體)和p75(Ngf、Bdnf、Nt3、Nt4/5和Nt6/7的通用神經(jīng)營養(yǎng)因子受體)。這一方面尤其具有吸引力,因?yàn)樗赡鼙砻鳎赽dnf缺失的遺傳背景下,神經(jīng)元存活途徑(即Trk2a和Trk3a的上調(diào))與死亡途徑(即p75的上調(diào))在發(fā)育過程中發(fā)生了必要的再平衡。為了深入了解差異表達(dá)基因的功能,我們對上調(diào)和下調(diào)基因進(jìn)行了基因本體論 (GO) 富集分析。我們確定了與bdnf通路相關(guān)的幾個(gè)類別,如神經(jīng)系統(tǒng)發(fā)育和功能、光感受、游泳行為,以及有趣的生物鐘。這些結(jié)果證實(shí)了bdnf突變系誘導(dǎo)轉(zhuǎn)錄組水平上的生物學(xué)變化。
Bdnf 調(diào)節(jié)晝夜節(jié)律:為了驗(yàn)證轉(zhuǎn)錄組學(xué)分析的結(jié)果,我們研究了8 dpf 的bdnf+/+和bdnf?/?仔魚在12:12明暗(LD)周期或恒定黑暗(DD)條件下時(shí)鐘和時(shí)鐘相關(guān)基因的表達(dá)模式。在突變仔魚中,ZT9處arntl1a表達(dá)水平的增加被顯著抑制。在DD條件下,正如預(yù)期的那樣,bdnf+/+仔魚保持了時(shí)鐘控制基因表達(dá)的節(jié)律模式,并顯示出光誘導(dǎo)基因per2和cry1a mRNA的顯著抑制。相反, bdnf ?/?仔魚, clock1a和clock2的表達(dá)譜在24小時(shí)內(nèi)出現(xiàn)失常。參與褪黑激素合成和調(diào)節(jié)的時(shí)鐘控制基因在魚類晝夜節(jié)律計(jì)時(shí)系統(tǒng)中也發(fā)揮著重要作用。aanat2是松果體中催化褪黑素合成的酶,其轉(zhuǎn)錄水平在光照期開始時(shí)較低,然后升高,在夜間(ZT21)結(jié)束時(shí)達(dá)到峰值。

圖4. 斑馬魚仔魚生物鐘基因的晝夜表達(dá)水平

圖5、斑馬魚仔魚生物鐘控制基因的晝夜表達(dá)水平
討論:在這項(xiàng)研究中,我們研究了Bdnf在斑馬魚發(fā)育過程中的作用,通過使用專門編輯目標(biāo)基因的最具創(chuàng)新性的技術(shù)CRISPR/Cas9系統(tǒng)產(chǎn)生40 bp敲除。在確認(rèn) bdnf 外顯子 2 中的選定突變導(dǎo)致 Bdnf 丟失后,我們分析了與 bdnf-/- 突變體相關(guān)的轉(zhuǎn)錄和行為表型,揭示了參與調(diào)節(jié)晝夜節(jié)律的關(guān)鍵基因的損傷。目前,關(guān)于Bdnf在晝夜節(jié)律調(diào)節(jié)中的作用的數(shù)據(jù)很少。本工作中描述的實(shí)驗(yàn)使驗(yàn)證轉(zhuǎn)錄組分析的結(jié)果成為可能,因?yàn)椴煌獯碳l件(即 LD 或 DD 周期)下的行為測試證實(shí)了晝夜節(jié)律的改變。22hpf的bdnf?/?仔魚經(jīng)7,8-DHF處理發(fā)現(xiàn)當(dāng)進(jìn)行運(yùn)動分析測試時(shí),拯救部分突變表型,從而證實(shí)突變行為實(shí)際上是由于缺乏Bdnf蛋白。在動物模型中進(jìn)行的體內(nèi)實(shí)驗(yàn)再現(xiàn)了人類疾病的特定特征,為理解該疾病的發(fā)育起源提供了機(jī)會。在神經(jīng)發(fā)育背景下,斑馬魚代表了一種強(qiáng)大的生物學(xué)工具,用于研究與遺傳畸變相關(guān)的特定機(jī)制。在動物模型中模擬突變?nèi)绾斡绊懮窠?jīng)系統(tǒng)發(fā)育,對于突出抑郁癥和焦慮等神經(jīng)疾病的潛在機(jī)制至關(guān)重要,這些疾病與Bdnf缺乏或晝夜節(jié)律改變密切相關(guān)。因?yàn)閎dnf在脊椎動物中高度保守,推導(dǎo)出斑馬魚氨基酸序列與哺乳動物的同源基因91%相似,我們假設(shè)在我們的突變魚類中觀察到的分子和行為表型可能會進(jìn)行臨床轉(zhuǎn)化。
以前的研究報(bào)道,小鼠bdnf突變體表現(xiàn)出負(fù)責(zé)運(yùn)動、平衡和姿勢控制的前庭神經(jīng)節(jié)神經(jīng)元減少。在小鼠模型中,闡明BDNF作用的一個(gè)主要障礙是純合突變體在出生后的早期致死性,通常發(fā)生在出生后2天內(nèi);只有一小部分動物最多能活2-3周。到目前為止,所有可用的結(jié)果都來自于對雜合子動物的研究,這些雜合子動物已達(dá)到成年?duì)顟B(tài),并且看起來有生育能力。由于少量的BDNF可能能夠發(fā)揮其自身的功能,因此完全敲除BDNF對于充分了解這種神經(jīng)營養(yǎng)素發(fā)揮的許多生物學(xué)功能至關(guān)重要。構(gòu)建的bdnf-/- 斑馬魚品系可以幫助彌補(bǔ)這一差距并提供新的見解。盡管我們對其生理學(xué)仍然知之甚少,但這里介紹的斑馬魚突變系能夠存活到成年期,并且可能解釋來自旁系同源基因所施加的“代償網(wǎng)絡(luò)效應(yīng)”的激活,這些基因已被證明可以緩沖基因組編輯誘導(dǎo)的有害突變 。在 bdnf+/- 小鼠中,BDNF 在 SCN 中的顯著抑制表達(dá)和 DD 中光誘導(dǎo)的運(yùn)動活動表明這種神經(jīng)營養(yǎng)因子在調(diào)節(jié)光夾帶中起作用。在將外源性BDNF注入SCN改變對光脈沖的行為反應(yīng)的大鼠試驗(yàn)中也獲得了類似的結(jié)果。由于這一關(guān)鍵問題之前尚未研究過,我們還研究了Bdnf在斑馬魚生物鐘系統(tǒng)中的作用。
事實(shí)上,在突變體中,DD條件下的運(yùn)動活動是失常的,并且通過藥物治療部分恢復(fù)了節(jié)律行為,證明了該模型的有效性。此外,DD 突變仔魚中 mRNA 表達(dá)模式的差異表明 Bdnf 對 clock1a 和 clock2(晝夜節(jié)律反饋回路的兩個(gè)正元件)以及編碼催化褪黑激素合成的酶 aanat2 的調(diào)節(jié)作用。魚類生物鐘的一個(gè)要點(diǎn)是松果體有節(jié)奏地合成和分泌褪黑素激素。褪黑激素在夜間產(chǎn)生,其產(chǎn)生速度取決于 AANAT 的酶活性。夜間高褪黑激素水平反映了 AANAT 合成和活性的增加,而白天褪黑激素產(chǎn)生的終止反映了這種酶的蛋白酶體降解。總體而言,這些數(shù)據(jù)首次在 bdnf-/- 斑馬魚仔魚中收集,為將 Bdnf 在晝夜節(jié)律中的多效性提供了一個(gè)新框架。 然而,需要進(jìn)一步的實(shí)驗(yàn)來確定可以直接或間接將 Bdnf 連接到分子生物鐘網(wǎng)絡(luò)的中間因素。靶向Bdnf-TrkB通路下游的特定Bdnf信號成分對于揭示這一有趣分子的復(fù)雜作用是必要的。
總之,我們描述了第一個(gè)可行的 bdnf-/- 脊椎動物模型,該模型為神經(jīng)營養(yǎng)因子的分子和細(xì)胞機(jī)制提供了獨(dú)特的見解。我們證明斑馬魚幼體中的Bdnf對行為晝夜節(jié)律的產(chǎn)生至關(guān)重要,并能更好地進(jìn)行人類神經(jīng)發(fā)育和晝夜節(jié)律調(diào)節(jié)疾病臨床轉(zhuǎn)化。這項(xiàng)研究證實(shí)了 Bdnf 參與的眾多途徑,并且構(gòu)建的斑馬魚突變系可作為未來研究的獨(dú)特工具,旨在了解 Bdnf 在不同生物過程中的作用,如神經(jīng)元發(fā)育和突觸傳遞、視覺光感知、 再生和新陳代謝。
原文出自:Loss of circadian rhythmicity in bdnf knockout zebrafish larvae - ScienceDirect
| 上一篇:魚類體內(nèi)新型污染物值得關(guān)注 | 下一篇:Molecular Biology and Evolution:穴居魚類神經(jīng)和全身脂質(zhì)代謝途徑的洞穴適應(yīng)研究取得進(jìn)展 |
|
|
|
在線 技術(shù)在線服務(wù)時(shí)間:9:00-20:00 在美鳳力,您對接的是技術(shù)人員,而非客服傳話!
電話
17312606166
微信
加微信獲取對接
|
