

TEL:17312606166(魏經理)



美鳳力臨床前大動物實驗中心

 17312606166
17312606166
感知機械信號的能力是生物體與周圍環境相互作用的基礎,對于生物體的生存至關重要。機械感受神經元通過將外界的機械刺激轉化為胞內信號,從而開啟感受神經通路。為了完成這一任務,神經元發育出了特化的亞細胞結構——力感受器(MO),它是由細胞膜上的力敏感通道和配套的結構組分(如細胞骨架以及細胞外基質)構成的一種“結構-力學”整體。在這一結構中,力敏感離子通道是信號轉導的關鍵,然而其結構組分則決定了機械感受神經元的感覺特性(如敏感性、響應范圍等)。為了匹配神經元的最佳感受模式,力感受器形成了獨有的特化結構(圖一),這些結構也構成了機械力信號轉導的“結構-力學”基礎。因此,探究力感受器結構的組成和發生,對于理解機械力信號的轉導機制以及相關的結構發育具有重要意義。
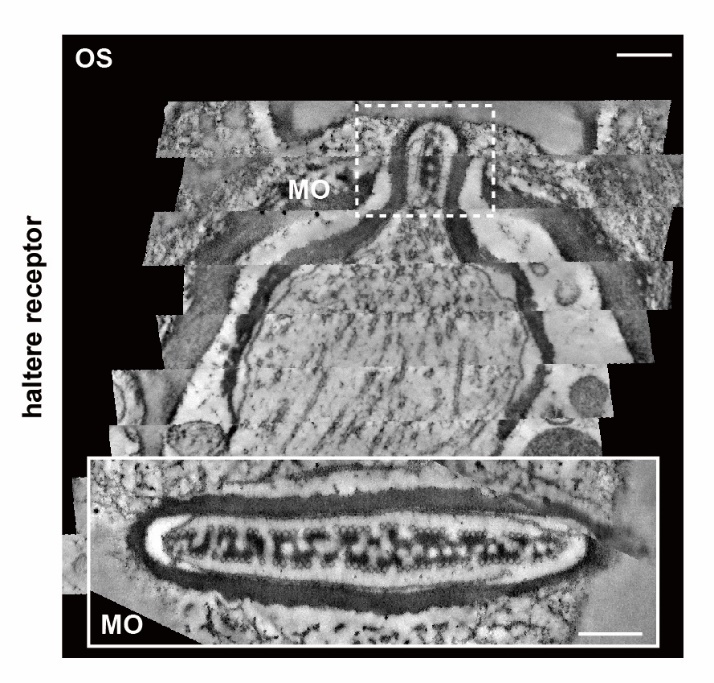
圖一.果蠅力感受器的超微結構
清華大學生命科學學院、清華-IDG/麥戈文腦科學研究中心梁鑫課題組在《細胞生物學雜志》(Journal of Cell Biology)期刊在線發表了題為“DCX-EMAP是果蠅力感受器超微結構的核心組織者” (DCX-EMAP is A Core Organizer for the Ultrastructure of Drosophila Mechanosensory Organelles)的研究論文。該研究通過解析果蠅力感受器的三維超微結構組織,明確了力感受器的主要結構復合細胞骨架是由短微管陣列與沉積在微管壁上的電子致密物質(EDM)共同組成。同時,該研究發現一種特異定位于力感受器的微管相關蛋白DCX-EMAP是果蠅機械力信號轉導所必需的。DCX-EMAP的缺失會導致果蠅力感受器中的微管數量顯著降低,微管陣列嚴重受損,并伴有電子致密物質的大量消失,表明DCX-EMAP在力感受器超微結構的組織中發揮了重要作用。
研究人員進一步揭示了DCX-EMAP在分子水平上的作用機制,發現該蛋白在力感受器的組織過程中具有雙重作用:一方面其氨基端的DCX結構域以抑制微管末端(包括生長末端及回縮末端)微管蛋白二聚體的解離發揮穩定微管的作用;而其羧基端的HELP以及WD40結構域則決定了DCX-EMAP在力感受器的特異性定位,其中WD40結構域還參與在局部空間中組織電子致密物質形成有序結構的過程。因此,DCX-EMAP通過同時整合微管與電子致密物質,在果蠅力感受器超微結構的組織過程中發揮了核心組織者的作用。該項工作以DCX-EMAP為入手點,系統地闡述了果蠅力感受器結構組織的分子機制,為理解機械力信號轉導分子機器的發生提供了新的見解。同時,對DCX-EMAP分子機制的研究,也擴展了對于DCX家族蛋白在纖毛中作用的認識,而這些發現也為纖毛病的治療提供了重要參考。
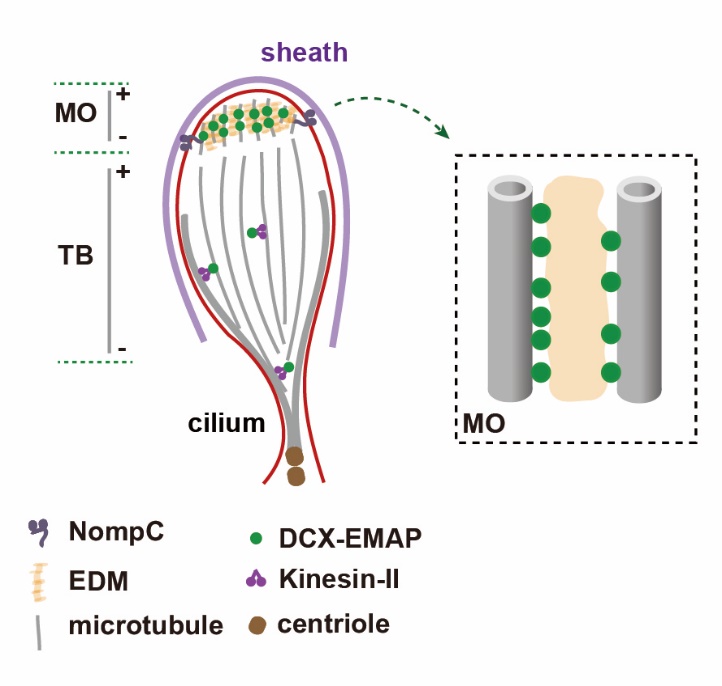
圖二.DCX-EMAP參與力感受器結構組織的分子機制
梁鑫課題組2018級博士生宋雪薇為本文第一作者,博士后崔立虹、2018級博士生王珊及多位實驗室同學和同事為實驗進行提供了重要幫助。梁鑫副教授為本文通訊作者,已出站博士后孫蘭弟(現任廣州實驗室高級工程師)為共同通訊作者。本研究得到了國家自然科學基金、科技部重點研發計劃及清華-IDG/麥戈文腦科學研究院的支持。
| 上一篇:許曉陽/章雪晴團隊開發mRNA-LNP介導的細胞焦亡療法,增強腫瘤免疫治療效果 | 下一篇:測量線粒體DNA損傷可預測帕金森病 |
|
|
|
|
